Optimal Weaning from an Evolutionary Perspective:
The evolution of our brains, meat eating, and a reliance on ketogenic metabolism
I recently had the privilege of presenting a talk, with the same title as this post, at the Ancestral Health Symposium.
I am posting the video here, with a transcript, some references, and related material.
Transcript
This is what I said for each slide, with comments / clarifications in square brackets. Times are approximate.
Optimal Weaning from an Evolutionary Perspective

0:00
My talk is called Optimal Weaning from an Evolutionary Perspective and I’d like to break down that title a little bit.
‘Optimal’ implies best for something, and here that something is going to be brain development.
The word ‘weaning’ can also benefit from clarification, because we often use it to mean the end of breastfeeding, but I use the convention meaning the beginning of the end, with the introduction of first foods.
For ‘evolutionary perspective’, I just want to point out that what we know about our past can inform our understanding of physiology, but our physiology can also constrain the possibilities of the past.
Overview

0:36
I’ve concluded that weaning infants onto an animal based diet best meets their nutritional needs, and the rest of this talk will be about why.
Primarily I’ll be talking about the unique properties that resulted from the evolution of our brains. I’ll also give a bit of evidence from modern health studies and trials, and then finally I’ll give a little bit of the how, based on my own experience in weaning one of my children onto animal based foods.
Human brains are unique

1:08
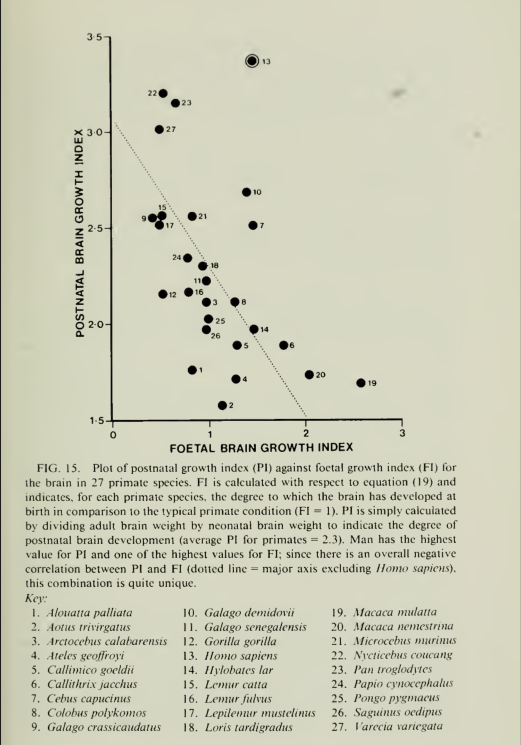
Human brains are unique in many ways, but one of the most striking things is their sheer size, especially relative to our bodies. In particular, when you take into account that we are primates, it’s really quite extraordinary. Primates already have brains that are about three times as large as most other mammals, at least relative to their size [1], and then humans have again about two and a half to three times as large brains as other primates do. And we didn’t always have that large a brain, that three times expansion occurred over the course of a few million years.
And a second related way that our brains are unique, is that our individual human brains do most of their growth after birth [2].
Altricial vs. Precocial

1:53
It’s helpful to think about this in context of the distinction between altricial and precocial animals, which is based on their degree of development at birth. Altricial animals are underdeveloped. They tend to have a short gestation, compared to precocial animals, who have a long gestation. They’re poorly developed, so they may be missing hair. They usually have underdeveloped sense organs, for example unopen eyes. They’re usually born in litters, as opposed to singletons, and they have less adult-like proportions, whereas precocial animals are essentially adult-like in their proportions. They have underdeveloped limbs, which means that they can’t do what precocial animals do,
which is move like the adults that they’re born from, and they tend to be smaller at birth, and their parents are younger when they reproduce.
Humans appear altricial but are precocial

2:41
Humans don’t really fit into this paradigm very well when you look at it at first glance, because they appear to be altricial, but they’re actually better understood as being precocial. Primates in general are highly precocial, and humans, when they fit that, are to the extreme, for example, we have enormous newborns, and we reproduce relatively late. Our babies appear altricial, though, because they’re born helpless, they don’t have adult proportions at all, and they can’t walk or have the motor skills that you would expect them to have.
But it’s helpful to think of them as actually precocial, but born early. And one reason to think that is because of fetal brain growth rates. We have our brains growing at the same rate as fetuses do persisting for up to a year [ should say at least ] after birth, and if you then look at our babies when they are a year old they look a lot more like you would expect them to look if they were born precocial: they have motor skills that you would expect them to have, and teeth, for example.
Human bains continue to grow postnatally
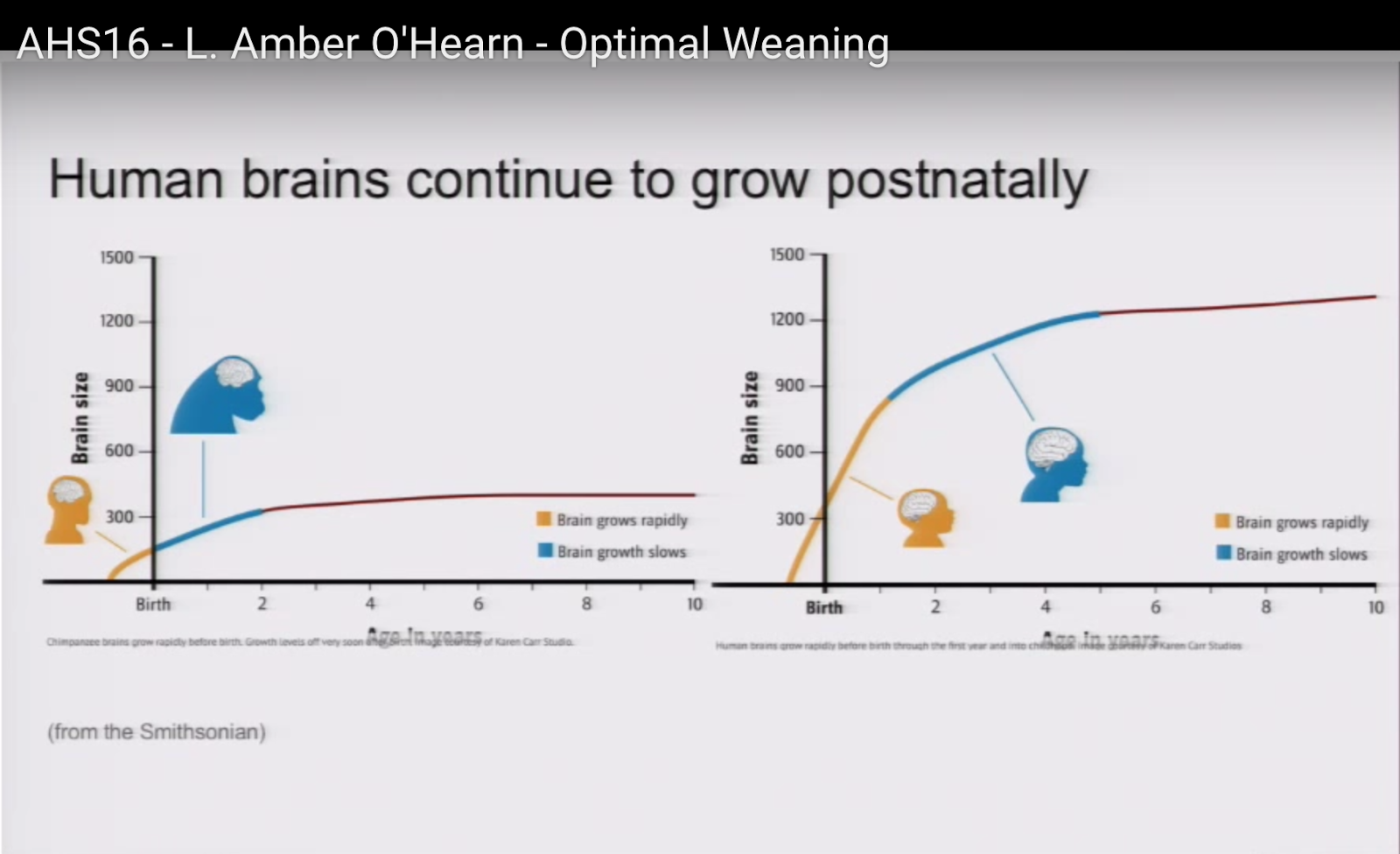
3:52
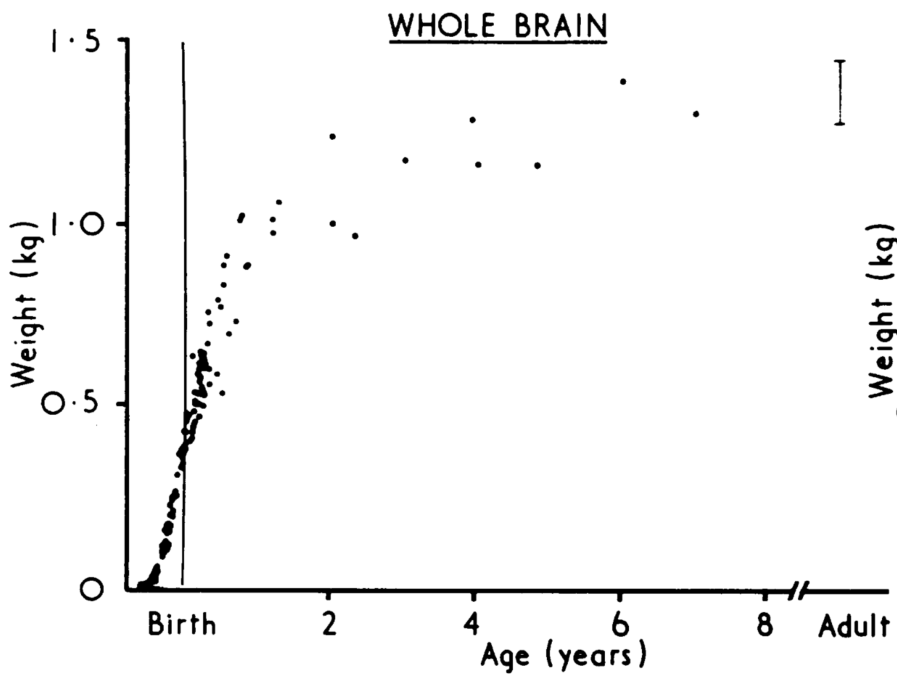
Here’s [sic] a couple of graphs from the Smithsonian. There’s one for chimpanzee brain growth and one for human brain growth. As you can see with the chimpanzee brain growth, they complete about half of their brain growth in gestation, and the rest over the course of a couple of years. Note that chimpanzees, like many primates wean quite late compared to us [3]; they wean at about four years of age, which is well after all their brain growth is completed.
Humans, on the other hand, have a very steep rate of growth before birth, and it continues into the second year [4], [5], and then the rate slows down some, although it’s still pretty significant, and then it’s followed by what looks on this graph like a levelling off, but this graph does end at age 10 and we know that there are growth spurts after that, too.
Rapid brain growth sustained beyond weaning

4:40
What I want to draw attention to with that is that the fetal-like brain growth doesn’t just extend beyond birth, but it also extends beyond the end of weaning. [This is a mistake. I meant to say that the rapid brain growth continues past the end of weaning, but it is “fetal-like” only past the beginning of weaning.]
So, we have this fetal-like growth in the first year, continued rapid growth to 5 years [ Or is it 4? ] [6], continued slower growth through childhood, and then, if you combine that with the fact that we wean early [3], we realise we need to support that kind of rate of growth even beyond weaning.
[ I’m also wondering whether weight is the best measure. Volume, density, cholesterol levels are all other measures to consider, but I won’t get into that here and now. ]
Our brains are really vulnerable, and they have many critical periods, each of which builds on the one before, so if you haven’t completed one of your stages of brain growth, you may not be able to complete the next stage successfully, and that means that you need continuous support through a long period of time [7].
Brain growth requirements

5:28
What kind of support do we need to give growing brains? Well there are at least three kinds of ways that we need to support a growing brain.
One is that they need specific micro-nutrients. Even adult brains can suffer if they don’t get enough of certain kinds of micronutrients and certainly developing brains that are missing these nutrients, if they’re missing them at critical times sometimes they can’t even recover from the detriment.
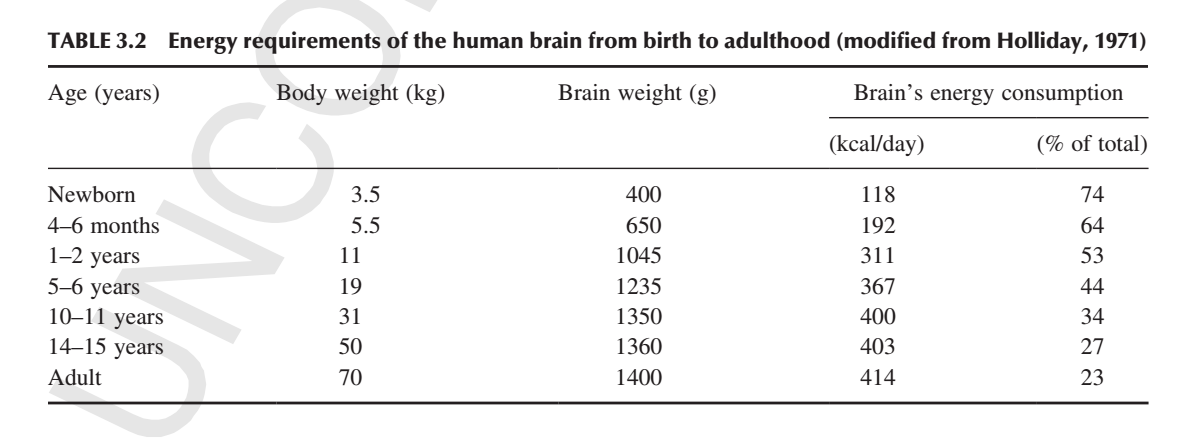
Secondly brains require an enormous amount of energy. At least 20% of the energy that we consume as adults goes to our brain and that’s even more extreme in a newborn who has about three quarters of the energy that they consume go[ing] right to the brain [8], [9].
And then thirdly, of course we need material for the structural components, and brains are made mostly of cholesterol and fat.
Brain evolution requirements

6:24
In parallel to that, the evolving of the brain has similar requirements. We needed those micronutrients and the energy and the structural components. We needed them to be available over a period of years for each individual and then that needed to be compounded more or less continuously for millions of years for us to be able to make that three times expansion.
Brain requirements: co-adaptations

6:51
While our brains were expanding over this long evolutionary period, there were co-adaptations that allowed them to expand, particularly contributing to the extraordinarily high energy requirements.
These co-adaptations I would like to talk about in more specifics: a high quality diet (by which I mean high in animal foods), shrinking intestines, a reliance on the ketogenic metabolism, and increased body fat particularly in babies.
Co-adaptation: eating meat
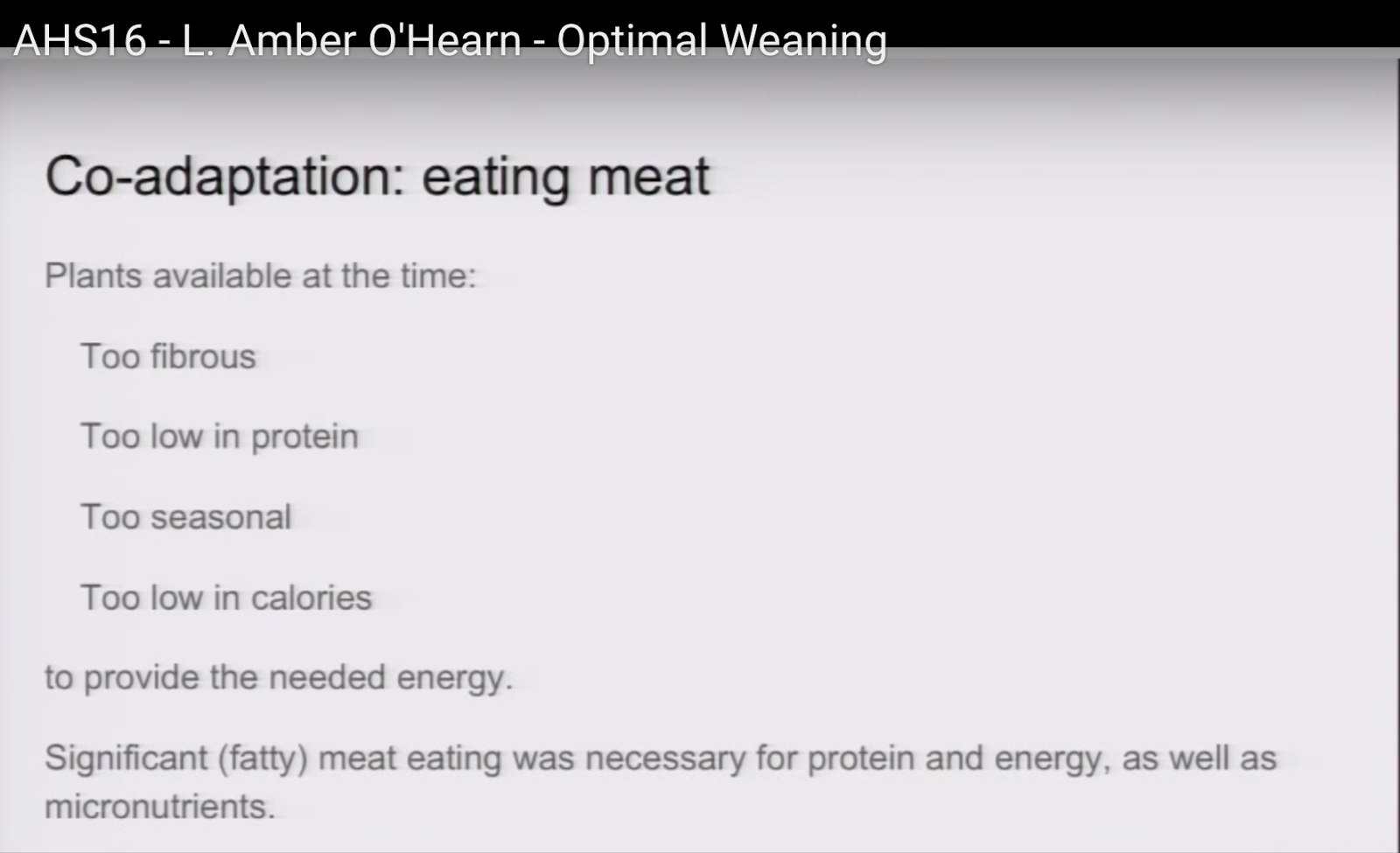
7:20
First of all, meat eating. The plants that were available to us at the time that we were expanding our brains were simply too fibrous, too low in protein, too seasonal, and too low in calories to provide the needed energy. So significant fatty meat eating was necessary for the protein and the energy as well as the micro-nutrients for developing our brains to our current form.
[ See the post, Meat is best for growing brains for more detail about the implausibility of plants as a sufficient food source.]
Brain requirements: micronutrients

7:39
I’m going to just zoom in on a few of those particularly critical micronutrients.
We have the minerals iodine [10] , iron [11], and zinc [12]; the fatty acid DHA [13], which is in all your brain cells in the phospholipids. It’s particularly important in vision (retinal cells) in the synapses, and vitamins A and D. If you don’t get enough of these vitamins and minerals and fatty acids as your brain is developing you can suffer developmental delay, disability. There is a tendency to emotional fragility and susceptibility to psychiatric disorders and it’s often not recoverable.
Micronutrient sources

8:27
For these micronutrients, animal foods are either the only, the best, or the most bioavailable source.
For DHA it’s almost exclusively found in animals. It’s true that there is some in microalgae, but it’s not very plausible but that’s where we were getting it while we were evolving.
Vitamin D is only available in animal sources. It’s true you can get it from sunshine, but again if you take into account the seasonality and the various geological periods we went through, It’s — we would need more.
Iron is available in plants, but it’s three times more bio-available in animal sources [14]. Similarly with vitamin A, which is 12 to 24 times more bioavailable in animal sources [15], [16]. If you think about the sheer amount of plant food that you would have to eat to try to make up for that, it’s just not plausible at all. For zinc it’s simply — animal sources are simply the best.
And then I’d also like to note that some plants actually interfere with the absorption of those minerals, so it might not just be not a benefit to try to get them from plants but it could actually be a detriment.
[ I refer the interested reader to the blog of Dr. Georgia Ede, and in particular, her post on vegetables ]
Co-adaptation: shrinking intestines

9:40
A second co-aptation is shrinking intestines. In 1995 Aiello and Wheeler came up with a hypothesis to try to explain how it could be that these brains that we’re growing which requires so much energy could have gotten that energy without giving up something else, and they noticed that we did give up something else. We gave up a drastic amount of the size of our intestines. Intestines are also really energy-intensive, so that smaller size freed up energy for the brain But there’s also a feedback loop, because having less intestines reduced our ability to consume fibre. A lot of other primates get a lot of their energy by consuming fibre and putting them through the factory of bacteria that turns that fibre into fat. We no longer have much of that ability at all and so that also increased our need to get our fat directly from an animal based diet.
[ See the post, Meat is best for growing brains for more detail about the the Expensive Tissue Hypothesis, and shrinking intestines. ]
Brain requirements: structural components

10:30
Going back to brain requirements, I wanted to re-emphasize the structural components I’d said that brains are mostly fat and cholesterol. By dry weight it’s about 60% lipids [17], about 40% of that which are cholesterol [18], but there’s a problem because fatty acids don’t cross the blood-brain barrier very easily [19], [ That is, DHA and AA enter the brain easily, but not the long chain fatty acids that white matter, gray matter, and myelin are mainly composed of. ] [20], [21] cholesterol almost not at all [22]. So all of that fat and cholesterol is reconstructed in the brain and it’s reconstructed we know, out of ketone bodies [ See next two slides ].
Ketone body fates
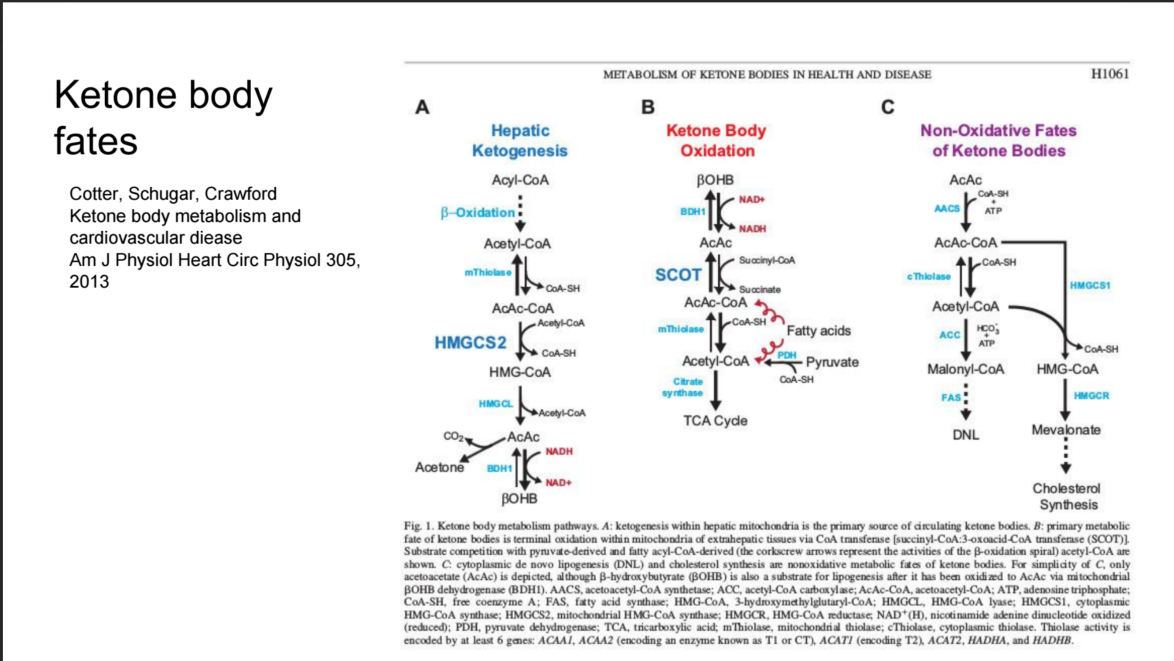
[ This slide I inadvertently omitted! It shows the biochemical pathways of ketone bodies being made in the liver, and what is relevant for this talk, being transformed into fuel, as is familiar to many, but also into fat and cholesterol, which may be new to many in audience. ]
Co-adaptation: reliance on ketogenic metabolism

11:00
That brings me to the third co-adaptation, which is using fat for energy and for substrates in the brain with ketone bodies. Ketone bodies are directly usable by the brain for energy, unlike fatty acids. They are used to create most of the fat and all the cholesterol.
[ Correction! Most of the fat and all of the cholesterol is synthesised in the brain, and preferentially by ketone bodies, but some is also made from glucose (which, of course, can be made on demand from protein. [23], [24], [25]]
and most importantly, they can easily and abundantly cross the blood-brain barrier.
There are other benefits to being in a ketogenic metabolism, for example, it increases the density of mitochondria in brain cells which allows more energy to flow and it also decreases the vulnerability of the growing brain to stress and trauma. You may be aware of the extreme neuroprotective properties of the ketogenic diet. For example, it mitigates drastically the damage that you would incur if you had a traumatic brain injury or stroke, so that’s obviously adaptive.
[ See the post The medical-grade diet, for more on neuroprotective properties of ketogenic diets. ]
Co-adaptation: reliance on ketogenic metabolism

11:52
The fact that we use ketone bodies for brain energy and material, which we and some other species also do in gestation, explains why newborns are in mild ketosis all the time [26]. Infants use ketones three to four times more efficiently than adults [ Correction! four to five times. (Three to four is in newborn rats.) ] [27], so mild ketonemia for a baby is more like a deeper ketosis for an adult. Even children as old as 12 and probably older can become ketogenic much more quickly and easily than you might expect. We’re talking about a matter of hours of fasting to develop the kind of ketosis that would take adults several days [28]. But even human adults become ketogenic more easily than other species and they do it without calorie restriction. This is really significant. I know of no other species that sustains ketosis without either starvation or semi-starvation, and it has implications for animal models of ketogenic diets therapies, because there may be cases where an animal requires caloric restriction for the therapy to be effective, whereas in humans it probably doesn’t, and would be a detriment to compliance and to health outcomes.
So I wanted to emphasize that humans have co-opted this trait that was previously an adaptation to cope with periods of starvation, and it still is in other species, but we have co-opted it into a default metabolism at least for the period of childhood to support the brain growth in particular, but also to meet the brain’s ongoing energy requirements.
Co-adaptation: increased body fat

13:30
Finally, the last co-adaptation I want to talk about is increased body fat, because it goes along with all the others It’s striking, again, when you compare humans with other primates, how fat they are. Even adults are fat compared to other primates. Other primates and most terrestrial animals actually have less than 5% body fat, and humans have easily somewhere between 15 and 20%, even very lean ones. Human babies take that to the extreme. They start out at about 15%. That’s doubled in a couple of months and it continues to increase over the first year.
Baby fat is different in character from the kind of fat you’d see in obese adults. It’s subcutaneous, not visceral [29], and it’s very low in polyunsaturated fatty acids even if their mother is eating a lot of polyunsaturated fatty acids, whereas obese adults tend to have a kind of roughly corresponding level and quality of fatty acids to what they’re eating. So there’s obviously a lot of filtering going on. And what polyunsaturated fatty acids are there are almost all DHA and arachidonic acid, which is another important brain fat, so it seems that this extreme body fat in babies is there to provide a continuous supply of fat that can be used by the brain both for energy and for materials via the ketogenic metabolism that we are relying on [30].
Summary of Evolutionary Evidence for Meat

[ This is the other slide I missed.]
14:56
I seem to be missing a slide.
I just wanted to quickly summarize what what I’ve said about evolution of the brain. The first is that we needed to evolve — we needed to eat meat to allow us to evolve the brains that we did. That’s for energy and for micronutrients. And I also wanted to emphasize the ketogenic metabolism part, because not only is it a natural normal default state for children but it shows that it’s not detrimental, it’s actually a benefit. It’s actually critical. It’s actually part of the mechanism of how we build our brains.
And so I’m bringing that up because someone who’s thinking about weaning their baby onto animal-based foods might worry: Wouldn’t this make them ketogenic and could that be a problem? And I just want to emphasize that not only is it not a problem, it’s the way it’s supposed to be and you could hardly stop if you wanted to because even when they sleep they’re going to go into ketosis.
Weaning onto meat: clinical trials

16:07
OK, so onto clinical trials. I know of two clinical trials that compared eating — weaning an infant onto the fortified cereals that we mostly recommend now, versus weaning them onto exclusively meat. The first one compared or took some measurements comparing them and the meat weaned children had a higher zinc status, which we know is very important. They had adequate iron without the benefit of supplementation that the cereal arm had. They had increased head growth which in children is a good index of brain growth, and it’s also correlated with higher intelligence and that’s not even taking into account the size of your head at birth so it’s not just the size of their head, it’s the amount of growth that happened between birth and the later time that’s correlated with the higher intelligence. And the second study just showed better general growth without increased adiposity That was what the researchers were worried about was that if you wean babies onto meat they would get fat in a way that would increase the risk for modern diseases and that of course didn’t happen.
Slide with refs

17:30
And I just have those references there for your reference. This kind of study is what I think has led to certain agencies like the Canadian government and the La Leche League to include meat as a recommended first food.
How? It’s easy.
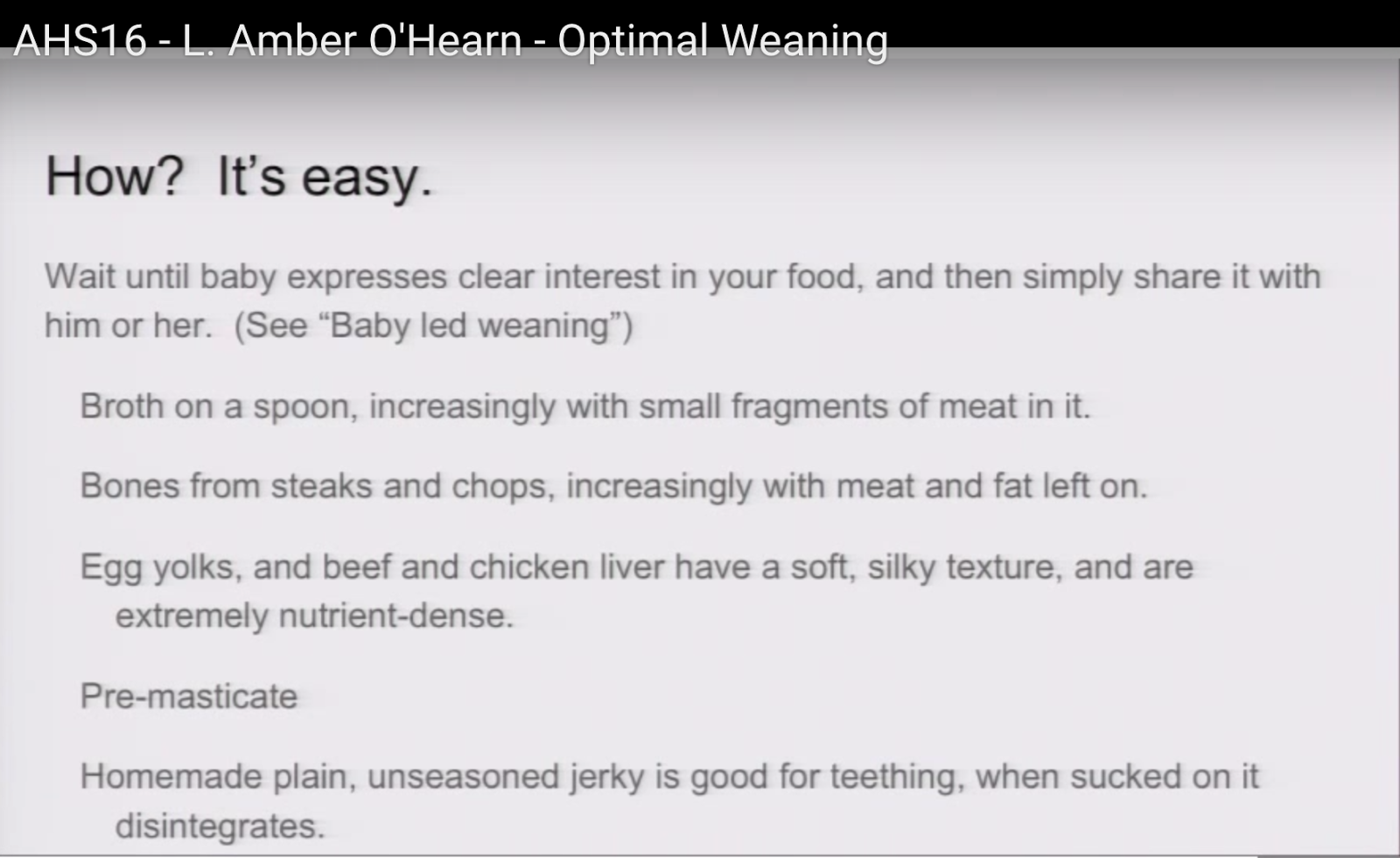
17:41
Finally I’m going to talk a little bit about how to do that just based on my experience from doing that with my third child. I was very influenced by Baby-Led Weaning. The core understanding from them is that you don’t — you can basically give a baby the same food that an adult eats. The risk of choking has been greatly exaggerated. You don’t need to buy into this whole, you know, factory-made baby food stuff. You can give them what you eat for the most part. So what I have done, for example:
I was in the habit of making bone broths that had some meat in the broth and I started by giving him broth on a spoon and increasingly over time added some fragments of meat.
I also gave him bones to teethe on from my steaks and chops, and again I increasingly left meat and fat on it, which he enjoyed a lot.
I fed him a lot of egg yolks and beef and chicken liver, which have a nice soft, silky texture. They’re extremely nutrient dense and to this day — this child is almost seven and liver is one of his favourite foods which pleases me to no end.
I’m really grateful to Aaron for being the first to bring up the word pre-masticate in this conference yesterday, so I didn’t have to be, and I also know from being in the audience that several people besides me did prechew their food for their babies and it’s certainly plausible — I would expect that a lot of people in the past did that and I did that.
I also often made plain unseasoned beef jerky which is really good for teething — sort of reminds me of a dog with rawhide he would gum down on it and pull and then he’d suck on it for a long time and it would basically just disintegrate. Also still one of his favorite foods.
(Photo slide)

19:35
And I’ll just leave you with a couple of photos of that baby who is almost 7. On the left here we have him at six months with a lamb bone that he was teething on. At the bottom: when he was two I discovered that he had liberated a stick of butter from the fridge, because that’s so delicious, and by two-and-a-half he was scrambling his own eggs.
This child basically had almost no plant matter in his diet for the first two years of his life and even now his diet is primarily animal-based.
Please give me your questions. Thank you.
Q&A
I’ve included the names of the questioners that I knew. If you are one I left out, introduce yourself!
Again, my clarifications or further comments in brackets.
Question 1 (Christopher Kelly)
C: I fully(?) subscribe to your ideas presented here and I have a very healthy two and a half year old daughter that’s eaten much the same way. But I thinks there’s an important point missing from your talk that is: the ketones come from medium chain triglycerides, that come from mom’s milk from eating carbohydrates. So the carbohydrates are synthesised in the breast tissue that make MCTs. Those MCTs are put in the breast milk, and that’s a really important ketogenic substrate, so I think that mom should be in ketosis, You see what I’m saying? The ketones should be synthesised by the baby.
A: I understand what you’re saying, yes. So I just want to give a couple of counter-examples. I didn’t eat any carbohydrate while I was making breast milk, and although there are medium chain triglycerides in the breast milk, that’s certainly not the only reason that babies are in ketosis. Even the babies postweaning that i mentioned earlier get into ketosis very rapidly, because it’s just the natural state. You can make it — that’s why what I’m positing here, and I’m not — it’s not my idea — but what I’m saying here is that the baby fat that is there is being turned into ketones just from the fat that’s stored on the body, just the same way that I make them
C: Right
A: And I forgot to mention that, I talked about how fat babies are and how fat adults are compared to other primates, and I think it’s quite significant. It would be unusual to see an animal that’s that fat if you thought that we weren’t naturally ketogenic animals.
C: Yeah and I’ve actually measured ketones, blood ketones, in my daughter when she was still an infant, breastfed only, and it was 1.6 mmol. But I can actually send you the studies that show that the the MCTs in milk, they go down and there’s studies where they’ve looked at giving MCTS to mom and they don’t go anywhere. Mom metabolizes them all. None of them appear in the breast milk, so I think like the carbohydrates for mom, It’s not my opinion, I can send you the studies that show that that might be important.
A: I’d like to see that. I guess what I’m trying to say is that the ketones that are in the baby’s blood don’t only come for a medium chain triglycerides…
C: Right, right, I understand that. Okay, yeah. I work with a doctor, he’s just finishing his PhD in neonatal neuroprotection so he’s done quite a lot of research in this area so I’ll send you some studies.
A: That’s fantastic. I would love that.
C: Okay, thank you.
Question 2
22:53
Q: Hey, that was great talk. Thank you. Can you say something about the timeframe and you’re, you know after three children and all of your research and interest in this, your thoughts on the timeframe for beginning the weaning process and then also how large that window of transition looks
A: Sure. There’s a lot I don’t know but I know that the recommendation currently is to start weaning at around four to six months and I think that the reason for that is because the amount of breast milk that children get, the caloric input just can’t provide much more than what they need by the time they’re that large and so I would say to start giving your baby food as soon as they start to
express interest in it. Just, you know, let them be the ones who say “I’m ready to start eating. Give me that.” And then how long it goes: Humans tend to wean a lot younger than other primates and I don’t know to what degree that’s enculturated and to what degree that’s natural. With my experience, my first child, I, he stopped breastfeeding at about two years and then each one after that was earlier and earlier with the last one, he stopped at nine months. So, I’m sorry I don’t know more about that.
Q: No, no, it’s okay. I just kind of wanted to see what your thoughts are. I suppose there’s some, aside from nutritional implications of how soon or early or late you you move away from breastfeeding I’m sure there’s other implications as well but it’s just it’s hard to understand. I just have a newborn, so I was just interested.
A: Congratulations!
Q: Yeah. Thank you so much!
Question 3 (Georgia Ede)
24:49
G: Amber, thank you for an exceptionally good talk. I just had a curiosity question as a psychiatrist. You having raised three children on this unique diet, which I wish were more common, [ Clarification: Unfortunately only my third child was weaning onto meat, though our household was always generally a low carb one. ] can you comment at all about how your children fared emotionally and physically compared to their peers? As a mother I would be very curious to hear.
A: Well there’s so much individuality I don’t want to necessarily claim too much. I know that my youngest child does have a very even temperament, especially compared to one of his brothers, but then on the other hand his oldest brother has perhaps the most even temperament of all, so don’t I don’t know what to conclude about that. One interesting thing that has been commented on to me many, many times is that my youngest child was never — he never missed a single day of daycare throughout — when he started at two and, so the entire three-year period, many of his peers, all of his peers missed significant time to many illnesses and he missed not a single day, so I like to attribute that to his diet.
G: Thank you very much. It was fascinating.
A: Thank you.
Question 4 (Ben Sima)
26:02
B: Has it been difficult to maintain his diet of high-fat from a social perspective, for example, they go over to someone else’s house and they have candy or something with other parents?
A: It is a challenge and the older they get the more of a challenge it is. My other children also at the time that I was weaning him, they were also transitioning to a more meat-based diet and yes it’s — I mean for example there, the number of special occasions that you have when you’re at school seem to be almost as numerous as the number of days Like, it’s always somebody’s birthday or some occasion and that’s always being celebrated with some kind of gluteny, sugary snack and yeah, it’s a struggle.
B: So do you find that he has a sweet tooth or does he kind of shun that
A: He loves sweet things when he gets his hands on them, but he doesn’t seem to be obsessed with them.
Question 5
27:04
Q: I just wanted to offer the cross cultural perspective that the worldwide age of breastfeeding cessation is about four to five years. [ But see footnote 3 below, which argues that the natural age is about 2.5 and for important, persuasive reasons. ] It’s only in the United States that it’s young, around a year, but if you look cross-culturally it is actually four years in most cultures.
A: Thank you for that. So that’s for the very end of breastfeeding?
Q: Yeah. So that’s kind of our biological norm. It’s more of a cultural thing here. The other thing that’s interesting is around four to six months — infants get a big bolus of iron from the placenta especially if we allow for delayed cord clamping — and then around four to six months that initial iron starts to go down which is another reason why, like you’re saying, meats are such a good first food, but that’s why that four to six months seems to be a good time to start foods is because that — not that breast milk is lacking in iron and zinc, but that that’s not where they’re supposed to get it from. You get for placentally and then it starts to go down around four to six months which is why, traditionally the idea of “ok let’s put iron in rice cereal” which we know — not a good idea but yeah that’s another reason why that four to six-month window seems to be a good time for getting those iron and zinc rich foods in.
Question 6 (Nick Mailer)
28:20
Q6: Thanks for the talk, it was very good. Something that hasn’t been discussed so much in this community is that weaning and continued breastfeeding is not merely about nutrition but it’s also about keeping the bond between the mother and the child, and that’s something that’s often overlooked. I know that there are people who I know who have generally weaned but, you know, when the child is a bit ill or is feeling a little bit insecure the child will revert for a little while to getting a little bit of breast milk or maybe once at night just to say goodnight. It becomes part of a ritual and part of a bonding process rather than as an essential continuing of nutrition, which is why as long as you’re comfortable with it there’s no harm in weaning in that extent, finally later, and that sometimes people feel the pressure — okay it’s four to six months I better stop by six months or something will happen — and people do feel that pressure which is why I think in the US and the UK people kind of feel that it’s a race to the final cessation of weaning and it doesn’t have to be as far as I’ve heard.
A: Right. Excellent point. Thank you.
Question 7
29:22
Hi. Thanks for the talk. So I have three children also. My youngest is 15 months. We thought a very similar, you know baby led weaning process, as your youngest. My question is, so my oldest is 11 too — quite a gap in between them, and you mentioned that vitamin D is one of the critical elements for brain development and prior to my son being born I had never seen like my pediatrician recommending vitamin D supplementation. So i guess my question is what are your thoughts on supplementing, like, drops as a newborn, and also what are some of the better animal sources other than I think fatty fish to get vitamin D from.
A: Yes, liver fatty fish… I’m surprised that you didn’t, weren’t recommended vitamin D drops because I remember that from even 15 years ago when my first son was born.
Q: Yeah I don’t remember if it’s possible. Five years between each of them. So, I think that’s weird you know.
A: Right.
Q: Did you do those?
A: I did. I did do those with the first two children. Actually I did it with all of them, come to think of it. Yeah.
Q: Thanks.
A: I figured there’s — the amount you would have to get to overdose is high enough that it wasn’t going to hurt.
Q: Yeah. We did it too. I just wasn’t sure. I hadn’t heard it before him, and you mentioned it, so thanks. Alright.
A: Thank you.
Question 8
30:53
Q: Hi. I missed the first part your talk which I’m bummed about, but I have a four-year-old who regularly steals butter out of the fridge, and her first foods were I think egg yolk, and I don’t think I did liver right away because I wasn’t doing that much liver but now she loves liver too. It’s like her favorite food.
A: Isn’t it good!
Q: Yeah, I mean I don’t particularly like it, but I eat it. But she like — she loves it.
I just wanted to add, too, maybe this will be covered in the next talk, about breastmilk and the microbiome, but one of the things that I found interesting about breastfeeding and the importance of it for the longer term is that it actually, the way that children remove milk from the breast actually helps to form the jaw and the palate, and so we see a lot today where women have to go back to work, you know six weeks, 12 weeks after giving birth and so they’repumping a lot and because we’re getting bottles and that’s really changing the way that our mouths are structured, I mean as are our nutrients in the womb and the palate formation.
I mean, anyone familiar with Weston Price’s work knows that, right, but I just think it’s an interesting piece, too, and I don’t think that there’s this — once kids start food they have to stop breast milk. In fact those things go together quite well for a long time because of the emotional factors and because of the palate formation and the muscle strength and the jaw formation.
A: Right.
Q: So, that I think is an interesting interesting piece, too, and yeah I’ve seen the same sort of statistics that hunter-gatherers usually breastfed three to four years, but they actually had a lower body fat, and so that would suppress ovulation for longer, which is why their children were spaced 4-5 years apart.
And there was no dairy. People weren’t eating dairy, so only dairy that was available was breast milk and the way that that dairy produces certain vitamins…
A: Lactose in particular is broken down into glucose and galactose and galactose is used to build some the brain material as well.
Q: There’s a question, so I have another question I’ll ask you later.
Question 9 (Kevin Boyd)
32:52
Q: Okay that was interesting. Who are you? That’s, it’s interesting that you’d, she’s — that’s my whole talk this afternoon. Please come.
Nutrition is concerned with nutrients, but not mechanical aspects of food processing and what she brought up was what I was gonna talk about a little bit, but how did you learn about baby led weaning, because, that is, for people who might not know could you explain a little bit about what it is and how you learned about it and how you are executing it with your own children?
A: Well, I’m not sure where I first heard of it, but the thing that I said that was the core important idea from it is what I’ve taken mostly from it, and that’s that babies don’t necessarily need you too mush everything up you can, you can give them a chicken drumstick and they’ll deal with it.
Q: Yeah. I’m going to really elaborate and so many wonderful points you made today, at one-thirty today.
A: Ok. Well, I won’t steal your thunder, then!
Q: It was a great talk.
A: Thank you.
Question 10
34:05
Q: Hi, I’m the token pre-mastication question. So you know, it goes: You know, you have your first baby and you sterilize everything before it touches her mouth and by the third baby you’re picking up a pacifier and you’re popping it in your own mouth before you pop it in theirs, and there was some concern about that in terms of, I guess, oral hygiene and what I had heard was, you know, it’s not such a wonderful thing to introduce your mouth germs to your baby, but if your pre-masticating their food perhaps you disagree with that.
A: Yes. Yes. I suppose if you had something unhealthy going on your mouth, that would be a problem, but I — I don’t really think that there’s anything unhygienic about the mouth, if you’re healthy.
Q: OK.
35:00
OK, well, thank you.
Acknowledgements
I’ve never given a talk to an audience of this size and calibre before. I particularly want to thank Sean Baker, Zooko Wilcox, and Jeff Pedelty for their support and encouragement in making it happen. I’m grateful also to the patient organisers of AHS for welcoming me and helping me through the process, particularly Katherine Morrison, Grace Liu, and Ben Sima.
References
Using EQ (encephalisation quotient), that is: a measure of relative brain size for mammals that takes into account some physical characteristics that affect the brain-body ratio.
2. Evidence type: experimental
Human brain evolution in an ecological context
Martin, Robert D.
Fifty-second James Arthur lecture on the evolution of the human brain 1982
3. Evidence type: review of data collection
From the ape’s dilemma to the weanling’s dilemma: early weaning and its evolutionary context
Kennedy GE1.
J Hum Evol. 2005 Feb;48(2):123-45. Epub 2005 Jan 18.
“Although humans have a longer period of infant dependency than other hominoids, human infants, in natural fertility societies, are weaned far earlier than any of the great apes: chimps and orangutans wean, on average, at about 5 and 7.7 years, respectively, while humans wean, on average, at about 2.5 years. Assuming that living great apes demonstrate the ancestral weaning pattern, modern humans display a derived pattern that requires explanation, particularly since earlier weaning may result in significant hazards for a child. Clearly, if selection had favored the survival of the child, humans would wean later like other hominoids; selection, then, favored some trait other than the child’s survival. It is argued here that our unique pattern of prolonged, early brain growth — the neurological basis for human intellectual ability — cannot be sustained much beyond one year by a human mother’s milk alone, and thus early weaning, when accompanied by supplementation with more nutritious adult foods, is vital to the ontogeny of our larger brain, despite the associated dangers.”
…
[On the data set:]
“Weaning is a process, not an event that can be placed at a specific point in time; therefore, it is not subject, in any meaningful way, to precise mathematical or statistical analyses or even to exact determination. Sellen’s (2001) recent paper has, perhaps, done as much as possible to overcome the inherent problems of determining human weaning time. An ‘‘average’’ age of weaning can only suggest the age at which most young in a particular group cease nursing; moreover, in humans, as the Amele demonstrate, it is not uncommon for a mother to continue to nurse an older youngster even though she has an infant as well. Data reported in Table 1 were taken from field studies, individual ethnographic reports, and from the Human Relations Area Files (HRAF: category 862, on-line edition); data points were included only when a definite age or clear range was expressed. All were pre-industrial, ‘‘natural fertility’’ populations practicing a range of subsistence economies from agriculture to foraging, and many were mixed economies.”
…
“Although a mean weaning age can be calculated from the human data in Table 1 (30.1 months; n = 46), it seems more accurate to conclude that the ‘‘natural’’ weaning age for humans is between 2-3 years and generally occurs about midway in that range. The minimum reported weaning age was one year (Fiji, Kogicol) and the maximum was about 4 years (several native American groups); several entries, however, reported that individual children may nurse as long as 6 years. Goodall (1986) also reported that a few Gombe chimps also nursed far longer than the population average. Sellen (2001), using a slightly larger sample (n = 113) also taken from the HRAF (microfiche edition), reported a very similar mean (29 months +/- 10 months), and a very similar peak weaning period between 2 and 3 years.”
…
“As noted below, stable nitrogen isotope analysis on bone tissue from several prehistoric societies suggests a somewhat wider range of ‘‘natural’’ weaning ages. For example, since nursing infants occupy a different (higher) trophic level than do their mothers, the isotopic composition of nursing infants’ bones and teeth should, in theory, differ from that of the adults in their group. Weaning time, therefore, should correspond to the point at which infant and adult tissues reach a similar isotopic composition ( Herring et al., 1998 ). Following Fogel et al. (1989), several authors have found an elevated level of δ15 N in infant osteological remains (relative to adults of the same group), which, they argued, constitutes a ‘‘nursing signal’’ ( Katzen- berg, 1992; Katzenberg et al., 1993, 1996; Schurr, 1994; Tuross and Fogel, 1994; White and Schwarcz, 1994 ). For example, at the Sully site in North Dakota and at the Angel site in the Ohio Valley, δ15 N reached adult levels at about 24 months (Tuross and Fogel, 1994; Schurr, 1997), suggesting rather early weaning. In Nubia, on the other hand, there was a gradual decline up to about age 6, indicating a slow introduction of adult foods ( White and Schwarcz, 1994 ). Others have used stable carbon and oxygen isotopes in dental enamel to track dietary changes in young children. Stable carbon (δ13 C), for example, may be used to detect the introduction of solid foods, and hence the beginning of the weaning period, while oxygen isotopes (δ18 O) may track the decreasing consumption of human milk (Wright and Schwarcz, 1998). Using this approach, it was found that, among the Preclassic and Postclassic Maya, solid foods were first introduced probably late in the first year, but that the weaning process was not concluded until 5 or 6 years ( Wright and Schwarcz, 1998).”
“[E]xtensive field data, collected in modern traditional societies, ancient textual references, and biochemical evidence from prehistoric societies, all suggest that in humans, the ‘‘natural’’ weaning age is generally between 2 and 3 years, although it may continue longer in some groups.”
4. Evidence type experiment
Quantitative growth and development of human brain
John Dobbing and Jean Sands
Arch Dis Child. 1973 Oct; 48(10): 757–767.
“One hundred and thirty-nine complete human brains ranging in age from 10 weeks’ gestation to 7 postnatal years, together with 9 adult brains, have been analysed in order to describe the human brain growth spurt quantitatively… The growth spurt period is much more postnatal than has formerly been supposed.”
[…]
“The postnatal cut-off point of the sigmoid curve of weight accumulation seems to be between 18 postnatal months and 2 years for whole brain.”
5. Evidence Type: review of experiments
Human brain evolution in an ecological context
Martin, Robert D.
Fifty-second James Arthur lecture on the evolution of the human brain 1982
[Emphasis ours]
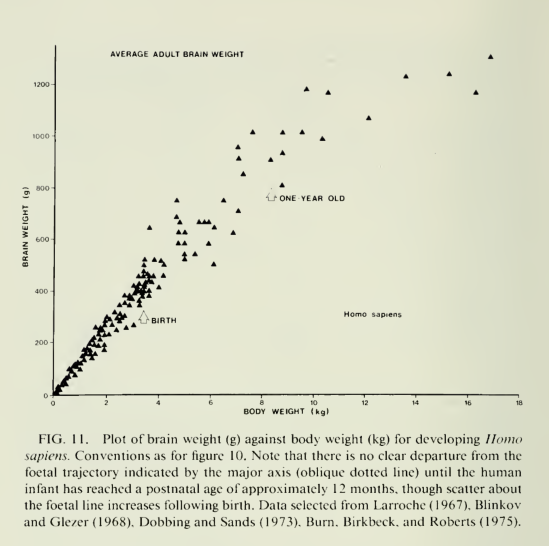
“The foregoing comparisons have demonstrated that Homo sapiens shares a number of general features of brain size and its development with the other primates, most notably in producing precocial off-spring and in the shift to a distinctive relationship between brain size and body size during foetal development (fig. 8). But human beings also exhibit a number of special features which set them apart from other primates, or at least from their closest relatives the great apes. These may be listed as follows:
- The remarkably large size of the adult brain relative to body size.
- The rapid development of both brain and body during foetal development, resulting in a distinctively large brain and body size at birth, compared to great apes.
- The greater degree of postnatal growth of the brain, accomplished by continuation of foetal brain : body relationships for at least one year after birth and associated with the “secondary altricial condition.“
[This shows pattern of brain to body weight ratio, not just brain weight]
6. Evidence type: experiment
Ann Neurol. 1978 Oct;4(4):345-56.
Changes in brain weights during the span of human life: relation of brain weights to body heights and body weights.
Dekaban AS.
“More than 20,000 autopsy reports from several general hospitals were surveyed for the purpose of selecting brains without a pathological lesion that had been weighed in the fresh condition. From this number, 2,773 males and 1,963 females were chosen for whom body weight, body height, and cause of death had been recorded. The data were segregated into 23 age groups ranging from birth to 86+ years and subjected to statistical evaluation. Overall, the brain weights in males were greater than in females by 9.8%. The largest increases in brain weights in both sexes occurred during the first 3 years of life, when the value quadruples over that at birth, while during the subsequent 15 years the brain weight barely quintuples over that at birth.”
[…]
“[T]he largest increases in brain weight occur during the first year of life, when the weight more than doubles that at birth (see Tables 2, 3; Figs 2A, 5). Further increases in brain weight also occur quite rapidly, although the increments from the preceding age groups are smaller. At about 3 years of age in males and between 3 and 4 years in females, the brain weight reaches four times the value at birth. Further growth of the brain is considerably slower, as it takes the brain 15 years (between ages 4 and 18) to nearly quintuple its birth weight and reach its mean highest value in young adults.”
7. Evidence Type: review of human and non-human animal experiments
Lipids, Malnutrition and the Developing Brain
CIBA Foundation Symposium
John Wiley & Sons, Sep 16, 2009 – Science – 192 pages
“The human growth spurt appears to extend throughout the last trimester of pregnancy and well into the 2nd year of postnatal life, and by analogy similar harm would be expected in the brains of humans growth-retarded during this time [as rats in reported nutrient deficiency experiment]
“There is no question but that the transient period of brain growth, known as the brain growth spurt, is more vulnerable to growth restriction than the periods both before and afterwards. Vulnerability in this sense means that quite mild restriction leads in experimental animals to permanent, irrecoverable reduction in the trajectory of bodily growth and to easily detectable distortions and deficits in the adult brain… In the present case the ‘damage’ consists of permanent but non-uniform reduction in the extent of brain growth. There is accumulating evidence that it has functional importance. An important feature of this type of vulnerability is that it is highly dependent on the timing of the insult, although not as finely so as the earlier teratology.”
8. Evidence type: review of experiments
Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition.
Siegel GJ, Agranoff BW, Albers RW, et al., editors.
Philadelphia: Lippincott-Raven; 1999.
“The brain consumes about one-fifth of total body oxygen utilization...
“The brain is metabolically one of the most active of all organs in the body. This consumption of O2 provides the energy required for its intense physicochemical activity. The most reliable data on cerebral metabolic rate have been obtained in humans. Cerebral O2 consumption in normal, conscious, young men is approximately 3.5 ml/100 g brain/min (Table 31-1); the rate is similar in young women. The rate of O2 consumption by an entire brain of average weight (1,400 g) is then about 49 ml O2/min. The magnitude of this rate can be appreciated more fully when it is compared with the metabolic rate of the whole body. An average man weighs 70 kg and consumes about 250 ml O2/min in the basal state. Therefore, the brain, which represents only about 2% of total body weight, accounts for 20% of the resting total body O2 consumption. In children, the brain takes up an even larger fraction, as much as 50% in the middle of the first decade of life [15].”
9. Evidence type: review
Human Brain Evolution: The Influence of Freshwater and Marine Food Resources
Stephen Cunnane (Editor), Kathlyn Stewart (Editor)
ISBN: 978-0-470-45268-4
June 2010, Wiley-Blackwell
10. Evidence type: review
The role of iodine in brain development
Delange F.
Proc Nutr Soc. 2000 Feb;59(1):75-9.
“I is required for the synthesis of thyroid hormones. These hormones, in turn, are required for brain development, which occurs during fetal and early postnatal life. The present paper reviews the impact of I deficiency (1) on thyroid function during pregnancy and in the neonate, and (2) on the intellectual development of infants and children. All extents of I deficiency (based on I intake (microgram/d); mild 50-99, moderate 20-49, severe > 20) affect the thyroid function of the mother and neonate, and the mental development of the child. The damage increases with the extent of the deficiency, with overt endemic cretinism as the severest consequence. This syndrome combines irreversible mental retardation, neurological damage and thyroid failure. Maternal hypothyroxinaemia during early pregnancy is a key factor in the development of the neurological damage in the cretin. Se deficiency superimposed on I deficiency partly prevents the neurological damage, but precipitates severe hypothyroidism in cretins. I deficiency results in a global loss of 10-15 intellectual quotient points at a population level, and constitutes the world’s greatest single cause of preventable brain damage and mental retardation.”
11. Evidence type: review
Micronutrient Deficiencies in the First Months of Life
edited by F. Delange, Keith P. West
“Iron plays a critical role in brain development, including its postnatal stages. Youdim et al., Youdim, Rouault and Beard reviewed the biological mechanisms whereby iron deficiency could possibly affect brain structure and functioning. The accumulation and distribution of iron in various regions of the brain depend on the stage of its development. This might indicate that brain regions vary in their vulnerability to iron deprivation, and suggests that the effect of iron deficiency on brain iron content could depend on the timing of the exposure. Animal studies indicate that low dietary intake of iron in the neonatal or preweaning period (before postnatal days 14-21 [ this must be rodents? ] may reduce whole-brain iron content that is not reversible by dietary repletion and produce irreversible effects. In rats, such effects occur before completion of brain organization and myelination and establishment of dopaminergic tracts. By contrast, dietary depletion in the postweaning period can also reduce brain iron content but this might be reversible upon dietary repletion. This illustrates that the timing of exposure to iron deficiency must be carefully considered when examining possible effects of iron deficiency on mental performance.
“Iron is not only required for brain growth and differentiation of neuronal cells, but also for protein synthesis, hormone production and other aspects of cellular enrgy metabolism and functioning. When sufficiently severe to reduce hemoglobin concentrations or cause anemia, iron deficiency might adversely affect oxygen delivery, thereby leading to reduced functioning of the central nervous system. Such deletrious effects of iron deficiency might be partially or completely reverese by iron repletion.
“Effects of iron deficiency might also be determined by other mechanisms. For example, it has been hypothesized that anemic children experience delayed acquisition of skills because they explore and interact less with their environment than nonanemic children, and they induce less stimulating behavior in their caretakers. Additionally, several studies have indicated that anemic children tend to be more fearful, withdrawn and tense, have reduced ability to focus their attention [25, 26], and are therefore less exposed to environmental stimuli that may promote mental and motor development.”
12. Evidence type: review (mostly non-human animal experiments)
Zinc and cognitive development.
Bhatnagar S, Taneja S.
Br J Nutr. 2001 May;85 Suppl 2:S139-45.
“Abstract
“Cognition is a field of thought processes by which an individual processes information through skills of perception, thinking, memory, learning and attention. Zinc deficiency may affect cognitive development by alterations in attention, activity, neuropsychological behavior and motor development. The exact mechanisms are not clear but it appears that zinc is essential for neurogenesis, neuronal migration, synaptogenesis and its deficiency could interfere with neurotransmission and subsequent neuropsychological behavior. Studies in animals show that zinc deficiency during the time of rapid brain growth, or during the juvenile and adolescent period affects cognitive development by decreasing activity, increasing emotional behavior, impairing memory and the capacity to learn. Evidence from human studies is limited. Low maternal intakes of zinc during pregnancy and lactation were found to be associated with less focused attention in neonates and decreased motor functions at 6 months of age. Zinc supplementation resulted in better motor development and more playfulness in low birth weight infants and increased vigorous and functional activity in infants and toddlers. In older school going children the data is controversial but there is some evidence of improved neuropsychological functions with zinc supplementation. Additional research is required to determine the exact biological mechanisms, the critical periods, the threshold of severity and the long-term effects of zinc deprivation on cognitive development.”
13. Evidence type: review
Role of omega-3 fatty acids in brain development and function: potential implications for the pathogenesis and prevention of psychopathology.
McNamara RK, Carlson SE.
Prostaglandins Leukot Essent Fatty Acids. 2006 Oct-Nov;75(4-5):329-49. Epub 2006 Sep 1.
“There is now good evidence suggesting that DHA is accrued in rodent, primate, and human brain during active periods of perinatal cortical maturation, and that DHA plays an important role in neuronal differentiation, synaptogenesis, and synaptic function. In animal studies, prenatal deficits in brain DHA accrual that are not corrected via postnatal dietary fortification are associated with enduring deficits in neuronal arborization, multiple indices of synaptic pathology, deficits in mesocorticolimbic dopamine neurotransmission, deficits in hippocampal serotonin and acetylcholine neurotransmission, neurocognitive deficits on hippocampus and frontal cortex-dependent learning tasks, and elevated behavioral indices of anxiety, aggression, and depression. Human and primate infants born preterm or fed diets without DHA postnatally exhibit lower cortical DHA accrual compared to infants born at term or fed human milk postnatally. Children/adolescents born preterm exhibit deficits in cortical gray matter expansion, neurocognitive deficits, and are at increased risk for attention-deficit/hyperactivity disorder (ADHD) and schizophrenia. Individuals diagnosed with ADHD or schizophrenia exhibit peripheral indices of lower DHA status and exhibit deficits in cortical gray matter expansion and deficits in cortical dopamine neurotransmission. Based on this body of evidence, it is hypothesized that perinatal deficits in brain DHA accrual represents a modifiable neurodevelopmental risk factor for the emergence of neurocognitive deficits and subsequent psychopathology. Evaluation of this hypothesis is currently feasible.”
14. Evidence type: review
Adaptation in iron metabolism.
J D Cook
Am J Clin Nutr February 1990 vol. 51 no. 2 301-308
[ Emphasis mine ]
“Dietary iron supply encompasses both the total amount of ingested iron and its bioavailability. Before 1950, nutritionists emphasized only total iron intake as a measure of dietary adequacy. Wider application of isotopic techniques during the l9SOs and l960s led to the realization that the bioavailability of ingested iron may be more important than total intake. There are two separate pathways of iron entry into the mucosal cell. The largest fraction of dietary iron is in the form of inorganic or nonheme iron, the absorption of which is determined largely by the nature of the meal. Nonheme-iron absorption occurs mainly from the duodenum because of the greater solubility of luminal iron in the proximal, more acid, region of the small intestine. Isotopic studies with extrinsic labeling demonstrated that all dietary forms of nonheme iron ingested in the same meal form a common pool within the intestinal lumen. Absorption from this pool is determined not by the type of the iron ingested but by enhancers, which promote absorption by maintaining iron in a reduced soluble form, and inhibitors, which bind iron or make iron insoluble and prevent its uptake by the brush border (29-32). The bioavailability of nonheme iron is enhanced by ascorbic acid and various tissue foods, such as meat, fish, and poultry, but not dairy products (33). A large number of dietary constituents impair iron absorption and these factors have been the major focus of absorption studies during the past decade. The most important inhibitors include tea, coffee, bran, calcium phosphate, egg yolk, polyphenols, and certain forms of dietary fiber. The extremes in bioavailability of nonheme iron as measured from isotopically labeled single meals served in a laboratory setting is nearly 15-fold (Fig 2). If tea is eliminated, absorption will increase about threefold. If meat is added, absorption will again increase 2-3 times. Maximal enhancement absorption occurs when a large quantity of ascorbic acid (eg, 2g) is taken with the meal.
“The second dietary iron fraction is heme, which is absorbed into the intestinal cell as an intact porphyrin complex. Specific receptors for heme iron have been identified in laboratory animals (34) but not in humans. After heme iron enters the cell it is rapidly degraded by heme oxygenase (35), and the released iron then enters the common intracellular iron pool. Subse- quent mucosal handling ofthis iron appears to be identical to that of inorganic iron. Because heme iron remains protected within the porphyrin complex before its uptake by the mucosa, it does not interact with dietary ligands and is therefore unaffected by the nature of the meal. Percentage absorption of heme iron is 5-10-fold higher than from nonheme iron. Although heme represents only 10-15% of dietary iron in meat-eating populations, it may account for nearly one-third of absorbed iron (36). Because absorption of heme iron is constant and independent of meal composition, the contribution of heme iron can be readily calculated from dietary records. This is in distinction to marked differences in the availability of non-heme iron.”
15. Evidence type: review
Consequences of Revised Estimates of Carotenoid Bioefficacy for Dietary Control of Vitamin A Deficiency in Developing Countries
Clive E. West*,†,2, Ans Eilander*, and Machteld van Lieshout*
J. Nutr. September 1, 2002 vol. 132 no. 9 2920S-2926S
“The bioefficacy of β-carotene in plant foods is much less than was previously thought. Intervention studies enrolled schoolchildren in Indonesia (10) and breast-feeding women in Vietnam (11) (Table 1). In each study there were four dietary groups: low-retinol, low-carotenoid diet (negative control); dark-green leafy vegetables (as well as carrots in the Indonesian study); yellow and orange fruits; and a retinol-containing diet (positive control). For dark-green leafy vegetables, the bioefficacy was 1:26 and 1:28; while for fruit, the bioefficacy was 1:12. This suggests that, with a mixture of vegetables and fruits in a ratio of 4:1, which is typical for both developing and developed countries, the bioefficacy of β-carotene from a mixed diet is 1:21. Chinese children aged 5–6.5 y yielded similar results for green and yellow vegetables (1:27) (14). Van Lieshout et al. (15), using the plateau isotopic enrichment method, also found relatively poor bioefficacy of β-carotene in dark-green leafy vegetables. β-Carotene in pumpkin was 1.7 times as potent as that in spinach (Table 1).”
See also the extensive review in the Vitamin A chapter of Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc Panel on Micronutrients, Subcommittees on Upper Reference Levels of Nutrients and of Interpretation and Use of Dietary Reference Intakes, and the Standing Committee on the Scientific Evaluation of Dietary Reference Intakes, which is accessible and would take a lot of space to include here.
17. Evidence type: review
The role of essential fatty acids in neural development: implications for perinatal nutrition.
Crawford MA.
Am J Clin Nutr. 1993 May;57(5 Suppl):703S-709S; discussion 709S-710S.
“The brain is 60% structural lipid, which universally uses arachidonic acid (AA; 20:4n6) and docosahexaenoic acid (DHA; 22:6n-3) for growth, function, and integrity. Both acids are consistent components of human milk. Experimental evidence in animals has demonstrated that the effect of essential fatty acid deficiency during early brain development is deleterious and permanent. The risk of neurodevelopmental disorder is highest in the very-low-birth-weight babies. Babies born of low birth weight or prematurely are most likely to have been born to mothers who were inadequately nourished, and the babies tend to be born with AA and DHA deficits. Because disorders of brain development can be permanent, proper provision should be made to protect the AA and DHA status of both term and preterm infants to ensure optimum conditions for the development of membrane-rich systems such as the brain, nervous, and vascular systems.”
18. Evidence type: experiment
Lipid composition of the normal human brain: gray matter, white matter, and myelin.
O’Brien JS, Sampson EL.
J Lipid Res. 1965 Oct;6(4):537-44.
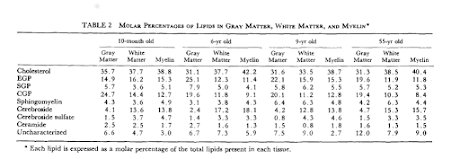
19. Evidence type: review
Blood-Brain Barrier Transport of Glucose, Free Fatty Acids, and Ketone Bodies
William M. Pardridge
Chapter in Fuel Homeostasis and the Nervous System, Volume 291 of the series Advances in Experimental Medicine and Biology pp 43-53
[ Emphasis mine ]
“Although free fatty acids are an important carbon source for cellular combustion in tissues such as skeletal muscle, fat, or liver in the postabsorptive state, brain does not significantly combust circulating free fatty acid, even after several weeks of prolonged starvation.(32) This failure to oxidize circulating free fatty acids is not due to a deficiency of the relevant free fatty acid oxidizing enzymes in brain since labeled free fatty acids are readily converted to CO 2 following the intracerebral administration of [14Cj-labeled free fatty acid,(33) and small amounts of circulating free fatty acids are converted to Krebs cycle intermediates (34). Rather, the failure of brain to utilize circulating free fatty acids as an important source of combustible carbon is due, in part, to a slow transport through the BBB. In the absence of plasma proteins, both medium chain and long chain free fatty acids are rapidly transported through the BBB. (35) However, free fatty acids are more than 99% bound by high affinity binding sites on circulating albumin, and only approximately 5% of plasma free fatty acid is unidirectionally extracted by brain on a single pass through the cerebral microcirculation .(36) Moreover, there is a prominent enzymatic barrier to the utilization of the circulating free fatty acids,3 as depicted in Figure 5. There is rapid esterification into membrane-bound triglyceride of circulating free fatty acid at either the endothelial membrane or the brain cell membrane. Thus, in the steady state, an equal amount of free fatty acid taken up by brain and esterified in the endothelial or brain cell membranes is released to blood via hydrolysis of membrane-bound triglyceride via brain microvascular lipoprotein lipase. 3 This enzymatic barrier protecting brain intracellular space from circulating free fatty acids is very well developed and breaks down only under pathologic conditions in brain.”
[ As far as I can tell, it has only recently been discovered that there are some mechanisms for transporting fatty acids across the blood brain barrier, but how much and under what circumstances is poorly understood. This statement expresses that candidly: ]
Blood-brain barrier and brain fatty acid uptake: Role of arachidonic acid and PGE2.
Murphy EJ.
J Neurochem. 2015 Dec;135(5):845-8. doi: 10.1111/jnc.13289. Epub 2015 Sep 17.
“How do fatty acids enter the brain and what role, if any, do membrane and cytosolic fatty acid binding proteins have on facilitating this process? This is a fundamental question that many lipid neurochemists will freely admit they cannot answer in any kind of definitive manner.”
20. Evidence type: experiment
Lipid composition of the normal human brain: gray matter, white matter, and myelin.
O’Brien JS, Sampson EL.
J Lipid Res. 1965 Oct;6(4):537-44.
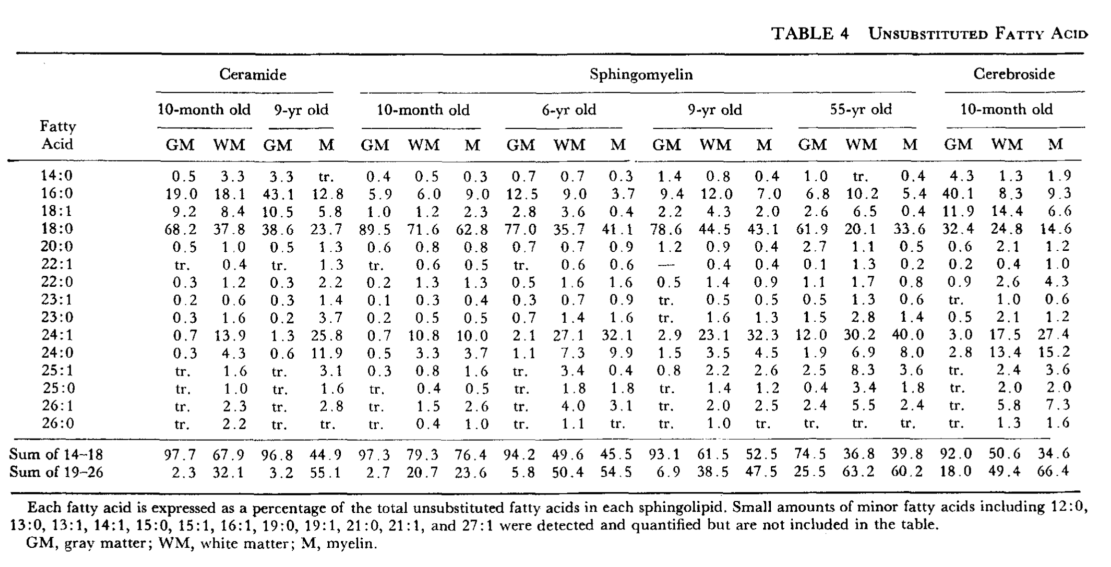
[ palimitic = 16:0, stearic = 18:0, oleic = 18:1 ]
21. Evidence type: non-human animal experiment
Fatty acid transport and utilization for the developing brain.
Edmond J, Higa TA, Korsak RA, Bergner EA, Lee WN.
J Neurochem. 1998 Mar;70(3):1227-34.
[ Emphasis mine ]
“To determine the transport and utilization of dietary saturated, monounsaturated, and n-6 and n-3 polyunsaturated fatty acids for the developing brain and other organs, artificially reared rat pups were fed a rat milk substitute containing the perdeuterated (each 97 atom% deuterium) fatty acids, i.e., palmitic, stearic, oleic, linoleic, and linolenic, from day 7 after birth to day 14 as previously described. Fatty acids in lipid extracts of the liver, lung, kidney, and brain were analyzed by gas chromatography-mass spectrometry to determine their content of each of the deuterated fatty acids. The uptake and metabolism of perdeuterated fatty acid lead to the appearance of three distinct groups of isotopomers: the intact perdeuterated, the newly synthesized (with recycled deuterium), and the natural unlabeled fatty acid. The quantification of these isotopomers permits the estimation of uptake and de novo synthesis of these fatty acids. Intact perdeuterated palmitic, stearic, and oleic acids from the diet were found in liver, lung, and kidney, but not in brain. By contrast, perdeuterated linoleic acid was found in all these organs. Isotopomers of fatty acid from de novo synthesis were observed in palmitic, oleic, and stearic acids in all tissues. The highest enrichment of isotopomers with recycled deuterium was found in the brain. The data indicate that, during the brain growth spurt and the prelude to myelination, the major saturated and monounsaturated fatty acids in brain lipids are exclusively produced locally by de novo biosynthesis. Consequently, the n-6 and n-3 polyunsaturated fatty acids must be transported and delivered to the brain by highly specific mechanisms.”
22. Evidence type: review
Brain Cholesterol: Long Secret Life Behind a Barrier
Ingemar Björkhem, Steve Meaney
Arteriosclerosis, Thrombosis, and Vascular Biology. 2004; 24: 806-815
“Although an immense knowledge has accumulated concerning regulation of cholesterol homeostasis in the body, this does not include the brain, where details are just emerging. Approximately 25% of the total amount of the cholesterol present in humans is localized to this organ, most of it present in myelin. Almost all brain cholesterol is a product of local synthesis, with the blood-brain barrier efficiently protecting it from exchange with lipoprotein cholesterol in the circulation. Thus, there is a highly efficient apolipoprotein-dependent recycling of cholesterol in the brain, with minimal losses to the circulation.Although an immense knowledge has accumulated concerning regulation of cholesterol homeostasis in the body, this does not include the brain, where details are just emerging. Approximately 25% of the total amount of the cholesterol present in humans is localized to this organ, most of it present in myelin. Almost all brain cholesterol is a product of local synthesis, with the blood-brain barrier efficiently protecting it from exchange with lipoprotein cholesterol in the circulation. Thus, there is a highly efficient apolipoprotein-dependent recycling of cholesterol in the brain, with minimal losses to the circulation.”
23. Evidence type: review
Cerebral ketone body metabolism. Morris AA
J Inherit Metab Dis. 2005;28(2):109-21.
“The second function of KBs in the brain is to provide substrates for the synthesis of various molecules. KBs are particularly important for the synthesis of lipids, such as cholesterol in myelin. Studies in 18-day-old rats found that KBs are incorporated into brain cholesterol and fatty acids much more readily than glucose is incorporated (Webber and Edmond 1977). Studies of cultured mouse astrocytes and neurons gave similar results (Lopes-Cardozo et al 1986). The preferential use of KBs for lipid synthesis probably occurs because they can be converted directly to acetoacetyl-CoA in the cytoplasm by acetoacetyl-CoA synthetase (EC 6.2.1.16, see Figure 1). Cytosolic acetoacetyl-CoA thiolase can then convert acetoacetyl-CoA to acetyl-CoA. Cytosolic acetyl-CoA can be generated from glucose (via the tricarboxylic acid cycle and ATP-citrate lyase, Figure 1) but this is a less direct pathway due to the inability of acetyl-CoA to cross the mitochondrial inner membrane. KBs are incorporated into fatty acids in the brain but they are primarily used for cholesterol synthesis (Koper et al 1981). Acetoacetyl-CoA synthetase expression in human brain parallels that of HMG-CoA reductase (EC 1.1.1.34), providing further evidence for the importance of this pathway in sterol synthesis (Ohgami et al 2003). Although KBs are the preferred substrates for brain lipogenesis, they appear not to be essential. Thus, rats fed a hypoketogenic diet develop normally (Auestad et al 1990). Development is also normal in most human patients with defects of ketogenesis (Morris et al 1998; van der Knaap et al 1998), though imaging sometimes shows white-matter abnormalities (see Clinical Considerations below).”
24. Evidence type: review
Preferential utilization of ketone bodies in the brain and lung of newborn rats.
Yeh YY, Sheehan PM.
Fed Proc. 1985 Apr;44(7):2352-8.
[ Emphasis mine ]
“Persistent mild hyperketonemia is a common finding in neonatal rats and human newborns, but the physiological significance of elevated plasma ketone concentrations remains poorly understood. Recent advances in ketone metabolism clearly indicate that these compounds serve as an indispensable source of energy for extrahepatic tissues, especially the brain and lung of developing rats. Another important function of ketone bodies is to provide acetoacetyl-CoA and acetyl-CoA for synthesis of cholesterol, fatty acids, and complex lipids. During the early postnatal period, acetoacetate (AcAc) and beta-hydroxybutyrate are preferred over glucose as substrates for synthesis of phospholipids and sphingolipids in accord with requirements for brain growth and myelination. Thus, during the first 2 wk of postnatal development, when the accumulation of cholesterol and phospholipids accelerates, the proportion of ketone bodies incorporated into these lipids increases. On the other hand, an increased proportion of ketone bodies is utilized for cerebroside synthesis during the period of active myelination. In the lung, AcAc serves better than glucose as a precursor forbiddingly the synthesis of lung phospholipids. The synthesized lipids, particularly dipalmityl phosphatidylcholine, are incorporated into surfactant, and thus have a potential role in supplying adequate surfactant lipids to maintain lung function during the early days of life. Our studies further demonstrate that ketone bodies and glucose could play complementary roles in the synthesis of lung lipids by providing fatty acid and glycerol moieties of phospholipids, respectively. The preferential selection of AcAc for lipid synthesis in brain, as well as lung, stems in part from the active cytoplasmic pathway for generation of acetyl-CoA and acetoacetyl-CoA from the ketone via the actions of cytoplasmic acetoacetyl-CoA synthetase and thiolase.”
25. Evidence type: non-human animal experiment
Acetoacetate and glucose as substrates for lipid synthesis by rat brain oligodendrocytes and astrocytes in serum-free culture.
Koper JW, Zeinstra EC, Lopes-Cardozo M, van Golde LM.
Biochim Biophys Acta. 1984 Oct 24;796(1):20-6.
“We have compared glucose and acetoacetate as precursors for lipogenesis and cholesterogenesis by oligodendrocytes and astrocytes, using mixed glial cultures enriched in oligodendrocytes. In order to differentiate between metabolic processes in oligodendrocytes and those in astrocytes, the other major cell type present in the mixed culture, we carried out parallel incubations with cultures from which the oligodendrocytes had been removed by treatment with anti-galactocerebroside serum and guinea-pig complement. The following results were obtained: 1. Both oligodendrocytes and astrocytes in culture actively utilize acetoacetate as a precursor for lipogenesis and cholesterogenesis. 2. In both cell types, the incorporation of acetoacetate into fatty acids and cholesterol exceeds that of glucose by a factor of 5-10 when the precursors are present at concentrations of 1 mM and higher. 3. Glucose stimulates acetoacetate incorporation into fatty acids and cholesterol, whereas acetoacetate reduces the entry of glucose into these lipids. This suggests that glucose is necessary for NADPH generation, but that otherwise the two precursors contribute to the same acetyl-CoA pool. 4. Both with acetoacetate and with glucose as precursor, oligodendrocytes are more active in cholesterol synthesis than astrocytes. 5. Using incorporation of 3H2O as an indicator for total lipid synthesis, we estimated that acetoacetate contributes one third of the acetyl groups and glucose one twentieth when saturating concentrations of both substrates are present.”
26. Evidence type: experiment
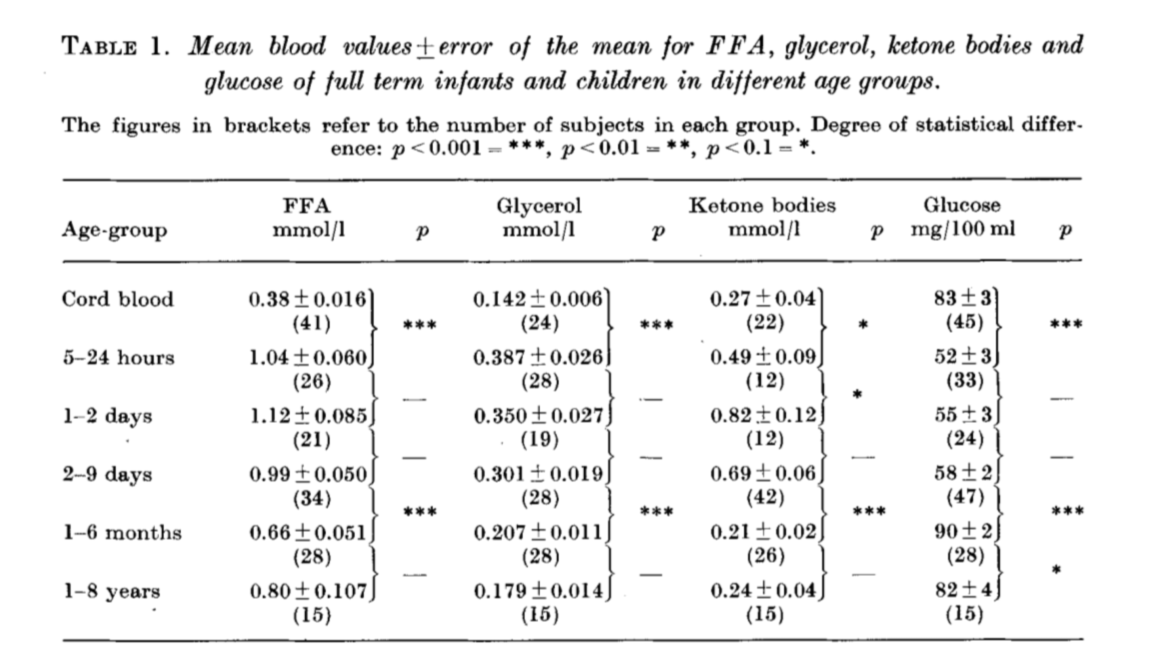
The pattern of blood lipids, glycerol and ketone bodies during neonatal period, infancy and childhood.
Persson B, Gentz J.
Acta Paediatr Scand 1966 Jul;55(4):353-62
“A total of 272 venous blood samples was obtained from umbilical cord and from children of varying ages from birth to 8 years. All were analysed for blood glucose and either FFA, glycerol or ketone bodies.”
[ Fasted overnight ]
27. Evidence type: experiment
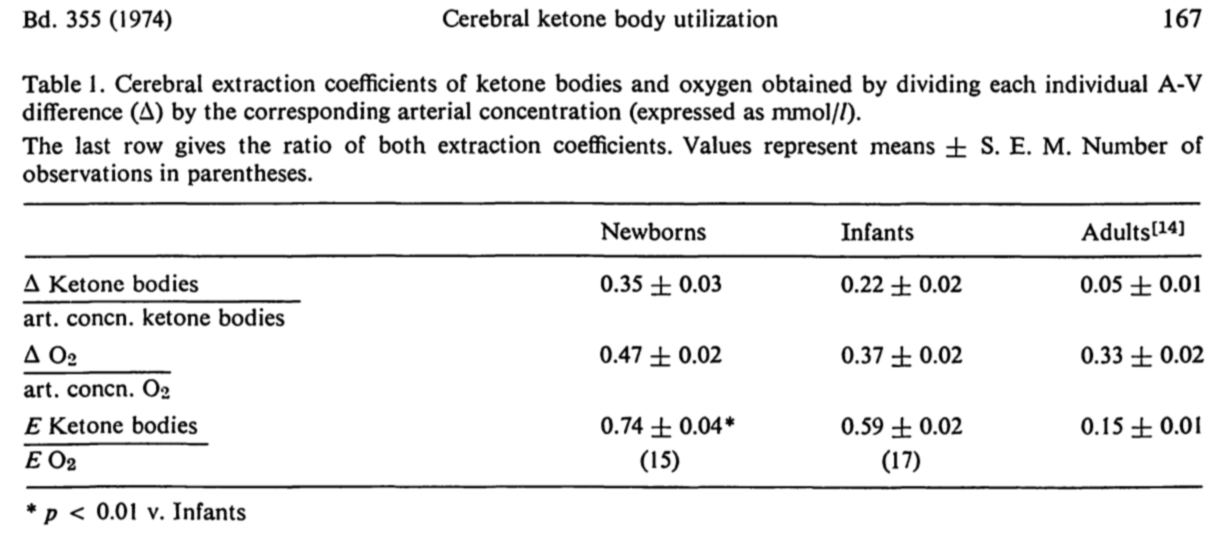
Developmental changes of cerebral ketone body utilization in human infants.
Kraus H, Schlenker S, Schwedesky D.
Hoppe Seylers Z Physiol Chem. 1974 Feb;355(2):164-70.
“Removal of circulating ketone bodies by the brain is greater in newborns than in infants. Both values are higher than those reported in adults [14]. This is demonstrated by the differences in the slopes of the’regression lines. From these data, however, the conclusion cannot be drawn that there is a specific enhancement of ketone body metabolism in the brains of young individuals. The total metabolic rate could be increased in the infant brain due to denser arrangement of blood capillaries, shorter diffusion distances and a higher cerebral blood flow. In order to avoid objections arising from these differences the contribution of ketone bodies to the total oxidative metabolism of the brain was calculated (last row of Table 1). Hence it follows that the brain’s capacity to utilize ketone bodies is specifically increased in newborns in comparison with infants. These values in turn are five and four times higher respectively than those reported in adults [14]. This conclusion is also justified by the finding that the contribution of glucose is not significantly altered throughout the different age groups. Corresponding relative values in newborns, infants and adults are 0.26, 0.27, and 0.33. The results of the present paper are confirmed by the report that the estimated cerebral uptake of ketone bodies in a group of older children (aged up to 14 years) was about three to four times higher than values observed in adults [19]. As cited above it was shown in different animals that the capacity to utilize ketone bodies is higher in the infant than in the adult brain [2, 20]. The increased ketone utilization by the animal brain during the neonatal period resulted from higher activities of the enzymes of ketone body utilization. Whether this also applies to the human infant brain remains to be tested.
28. Evidence type: experiment
Ketone body transport in the human neonate and infant.
P F Bougneres, C Lemmel, P Ferré, and D M Bier
J Clin Invest. 1986 Jan; 77(1): 42–48.
[ Emphasis ours ]
“Using a continuous intravenous infusion of D-(-)-3-hydroxy[4,4,4-2H3]butyrate tracer, we measured total ketone body transport in 12 infants: six newborns, four 1-6-mo-olds, one diabetic, and one hyperinsulinemic infant. Ketone body inflow-outflow transport (flux) averaged 17.3 +/- 1.4 mumol kg-1 min-1 in the neonates, a value not different from that of 20.6 +/- 0.9 mumol kg-1 min-1 measured in the older infants. This rate was accelerated to 32.2 mumol kg-1 min-1 in the diabetic and slowed to 5.0 mumol kg-1 min-1 in the hyperinsulinemic child. As in the adult, ketone turnover was directly proportional to free fatty acid and ketone body concentrations, while ketone clearance declined as the circulatory content of ketone bodies increased. Compared with the adult, however, ketone body turnover rates of 12.8-21.9 mumol kg-1 min-1 in newborns fasted for less than 8 h, and rates of 17.9-26.0 mumol kg-1 min-1 in older infants fasted for less than 10 h, were in a range found in adults only after several days of total fasting. If the bulk of transported ketone body fuels are oxidized in the infant as they are in the adult, ketone bodies could account for as much as 25% of the neonate’s basal energy requirements in the first several days of life. These studies demonstrate active ketogenesis and quantitatively important ketone body fuel transport in the human infant. Furthermore, the qualitatively similar relationships between the newborn and the adult relative to free fatty acid concentration and ketone inflow, and with regard to ketone concentration and clearance rate, suggest that intrahepatic and extrahepatic regulatory systems controlling ketone body metabolism are well established by early postnatal life in humans.”
29. Evidence type: experiment
Fast and reproducible method for the direct quantitation of adipose tissue in newborn infants.
Harrington TA, Thomas EL, Modi N, Frost G, Coutts GA, Bell JD.
Lipids. 2002 Jan;37(1):95-100.
“The role of body fat content and distribution in infants is becoming an area of increasing interest, especially as perception of its function appears to be rapidly evolving. Although a number of methods are available to estimate body fat content in adults, many are of limited use in infants, especially in the context of regional distribution and internal depots. In this study we developed and implemented a whole-body magnetic resonance imaging (MRI)-based protocol that allows fast and reproducible measurements of adipose tissue content in newborn infants, with an intra-observer variability of <2.4% and an inter-observed variability of <7%. The percentage total body fat for this cohort of infants ranged from 13.3-22.6% (mean and standard deviation: 16.6 +/- 2.9%), which agrees closely with published data. Subcutaneous fat accounted for just over 89% of the total body fat, whereas internal fat corresponded to almost 11%, most of which was nonabdominal fat. There were no gender differences in total or regional body fat content. These results show that whole-body MRI can be readily applied to the study of adipose tissue content and distribution in newborn infants. Furthermore, its noninvasive nature makes it an ideal method for longitudinal and interventional studies in newborn infants.”
30. Evidence type: review
Placental regulation of fatty acid delivery and its effect on fetal growth–a review.
Haggarty P
Placenta. 2002 Apr;23 Suppl A:S28-38.
[ Emphasis ours ]
“The likelihood that the composition of fatty acids delivered to the fetus can affect the quality of fetal development is more compelling. The concentration of DHA in the brain of neonates is dependent on the intake of pre-formed DHA (Farquharson et al., 1993; Jamieson et al., 1999; Makrides et al., 1994) and many workers have reported beneficial effects of LCPUFA intake in early post-natal life in particular (Birch et al., 1992 ; Hoffman et al., 1993; Horwood, Darlow and Mogridge, 2001; Lucas et al., 1992, 1998). In this context, much has been made of the relatively high concentration of DHA in the fetal brain at term and the importance of in utero DHA supply but this is not specifically a fetal/placental issue as the human brain, including that of the pregnant mother, maintains a high concentration of DHA throughout life. Furthermore the total amount of DHA present in the fetal brain at term is not much greater than that in the placenta itself. An issue that is much more clearly specific to the fetus and placenta is the very high concentration of DHA and AA achieved in the fetal adipose tissue and the fact that 16 times more DHA is stored in the adipose tissue than is deposited in the fetal brain during in utero life. Within a few hours of birth there is a dramatic rise in plasma TG and NEFA indicating mobilization of adipose tissue stores ( Van Duyne & Havel, 1959 ) such that the concentration of DHA in the adipose tissue is undetectable after two months of post-natal life on a diet devoid of pre-formed DHA ( Farquharson et al., 1993). Thus the importance of this adipose store of LCPUFA may be to protect processes such as brain and retinal development against a poor dietary supply of LCPUFA during the critical first months of post-natal life. The fact that most of the LCPUFA such as DHA which is accrued by the fetus is actually stored in fetal adipose tissue also implies that there is normally an excess availability in utero for development of the fetal organs and tissues and that it is only in low birth weight babies, where the body fat content may be very low (Sparks et al., 1980), that the supply of LCPUA may become limiting for fetal requirements during in utero life.”